Размножение бактерий путем деления — самый распространенный метод увеличения численности микробной популяции. После деления происходит рост бактерий до исходного размера, для чего необходимы определенные вещества (факторы роста).
Способы размножения бактерий различны, но для большинства их видов присуща форма бесполового размножения способом деления. Способом почкования бактерии размножаются исключительно редко. Половое размножение бактерий присутствует в примитивной форме.
Рис. 1. На фото бактериальная клетка в стадии деления.
Генетический аппарат бактерий
Генетический аппарат бактерий представлен единственной ДНК — хромосомой. ДНК замкнута в кольцо. Хромосома локализована в нуклеотиде, не имеющем мембраны. В бактериальной клетке имеются плазмиды.
Нуклеоид
Нуклеоид является аналогом ядра. Он расположен в центре клетки. В нем локализована ДНК — носитель наследственной информации в свернутом виде. Раскрученная ДНК достигает в длину 1 мм. Ядерное вещество бактериальной клетки не имеет мембраны, ядрышка и набора хромосом, не делится митозом. Перед делением нуклеотид удваивается. Во время деления число нуклеотидов увеличивается до 4-х.

Рис. 2. На фото бактериальная клетка на срезе. В центральной части виден нуклеотид.
Плазмиды
Плазмиды представляют собой автономные молекулы свернутые в кольцо двунитевой ДНК. Их масса значительно меньше массы нуклеотида. Несмотря на то, что в ДНК плазмид закодирована наследственная информация, они не являются жизненно важными и необходимыми для бактериальной клетки.

Рис. 3. На фото бактериальная плазмида.
Этапы деления
После достижения определенных размеров, присущих взрослой клетке, запускаются механизмы деления.
Репликация ДНК
Репликация ДНК предшествует клеточному делению. Мезосомы (складки цитоплазматической мембраны) удерживают ДНК до тех пор, пока процесс деления (репликации) не завершится.
Репликация ДНК осуществляется с помощью ферментов ДНК-полимеразами. При репликации водородные связи в 2-х спиральной ДНК разрываются, в результате чего из одной ДНК образуются две дочерние односпиральные. В последующем, когда дочерние ДНК заняли свое место в разделенных дочерних клетках, происходит их восстановление.
Как только репликация ДНК завершилась, в результате синтеза появляется перетяжка, разделяющая клетку пополам. Вначале делению подвергается нуклеотид, затем цитоплазма. Синтез клеточной стенки завершает деление.

Рис. 4. Схема деления бактериальной клетки.
Обмен участками ДНК
У сенной палочки процесс репликации ДНК завершается обменом участками 2-х ДНК.
После деления клетки образуется перемычка, по которой ДНК одной клетки переходит в другую. Далее обе ДНК сплетаются. Некоторые отрезки обоих ДНК слипаются. В местах слипания происходит обмен отрезками ДНК. Одна из ДНК по перемычке уходит обратно в первую клетку.

Рис. 5. Вариант обмена ДНК у сенной палочки.
Типы делений бактериальных клеток
Если клеточное деление опережает процесс разделения, то образуются многоклеточные палочки и кокки.
При синхронном клеточном делении образуются две полноценные дочерние клетки.
Если нуклеотид делится быстрее самой клетки, то образуются многонуклеотидные бактерии.
Способы разделения бактерий
Деление с помощью разламывания
Деление с помощью разламывания характерно для сибиреязвенных бацилл. В результате такого деления клетки переламываются в местах сочленения, разрывая цитоплазматические мостики. Далее отталкиваются друг от друга, образуя цепочки.
Скользящее разделение
При скользящем разделении после деления клетка обосабливается и как бы скользит по поверхности другой клетки. Данный способ разделения характерен для некоторых форм эшерихий.
Секущееся разделение
При секущемся разделении одна из разделившихся клеток свободным концом описывает дугу круга, центром которого является точка ее контакта с другой клеткой, образуя римскую пятерку или клинопись (коринебактерии дифтерии, листерии).

Рис. 6. На фото бактерии палочковидной формы, образующие цепочки (сибиреязвенные палочки).

Рис. 7. На фото скользящий способ разделения кишечных палочек.

Рис. 8. Секущийся способ разделения коринебактерий.
Вид скоплений бактерий после деления
Скопления делящихся клеток имеют разнообразную форму, которая зависит от направления плоскости деления.
Шаровидные бактерии располагаются по одному, по двое (диплококки), пакетами, цепочками или как гроздья винограда. Палочковидные бактерии — цепочками.
Спиралевидные бактерии — хаотично.

Рис. 9. На фото микрококки. Они круглые, гладкие, имеют белую, желтую и красную окраску. В природе микрококки распространены повсеместно. Живут в разных полостях человеческого организма.

Рис. 10. На фото бактерии диплококки — Streptococcus pneumoniae.

Рис. 11. На фото бактерии сарцины. Кокковидные бактерии соединяются в пакеты.

Рис. 12. На фото бактерии стрептококки (от греческого «стрептос» — цепочка). Располагаются цепочками. Являются возбудителями целого ряда заболеваний.

Рис. 13. На фото бактерии «золотистые» стафилококки. Располагаются, как «гроздья винограда». Скопления имеют золотистую окраску. Являются возбудителями целого ряда заболеваний.

Рис. 14. На фото извитые бактерии лептоспиры — возбудители многих заболеваний.

Рис. 15. На фото палочковидные бактерии рода Vibrio.
Скорость деления бактерий
Скорость деления бактерий крайне высока. В среднем одна бактериальная клетка делится каждые 20 минут. В течение только одних суток одна клетка образует 72 поколения потомства. Микобактерии туберкулеза делятся медленно. Весь процесс деления занимает у них около 14 часов.

Рис. 16. На фото отображен процесс деления клетки стрептококка.
Половое размножение бактерий
В 1946 году учеными было обнаружено половое размножение в примитивной форме. При этом гаметы (мужские и женские половые клетки) не образуются, однако некоторые клетки обмениваются генетическим материалом (генетическая рекомбинация ).
Передача генов осуществляется в результате конъюгации — однонаправленного переноса части генетической информации в виде плазмид при контакте бактериальных клеток.
Плазмиды представляют собой молекулы ДНК небольшого размера. Они не связаны с геномом хромосом и способны удваиваться автономно. В плазмидах содержаться гены, которые повышают устойчивость бактериальных клеток к неблагоприятным условиям внешней среды. Бактерии часто передают эти гены друг другу. Отмечается так же передача генной информации бактериям другого вида.
При отсутствии истинного полового процесса именно конъюгация играет огромную роль при обмене полезными признаками. Так передается способность бактерий проявлять лекарственную устойчивость. Для человечества особо опасным является передача устойчивости к антибиотикам между болезнетворными популяциями.

Рис. 17. На фото момент конъюгации двух кишечных палочек.
Фазы развития бактериальной популяции
При посевах на питательную среду развитие бактериальной популяции проходит несколько фаз.
Исходная фаза
Исходная фаза — это период от момента посева до их роста. В среднем исходная фаза длится 1 — 2 часа.
Фаза задержки размножения
Это фаза интенсивного роста бактерий. Ее длительность составляет около 2-х часов. Она зависит от возраста культуры, периода приспособления, качества питательной среды и др.
Логарифмическая фаза
В эту фазу отмечается пик скорости размножения и увеличения бактериальной популяции. Ее длительность составляет 5 — 6 часов.
Фаза отрицательного ускорения
В эту фазу отмечается спад скорости размножения, уменьшается количество делящихся и увеличивается число погибших бактерий. Причина отрицательного ускорения — истощение питательной среды. Ее длительность составляет около 2-х часов.
Стационарная фаза максимума
В стационарную фазу отмечается равное количество погибших и вновь образованных особей. Ее длительность составляет около 2-х часов.
Фаза ускорения гибели
В эту фазу прогрессивно нарастает количество погибших клеток. Ее длительность составляет около 3-х часов.
Фаза логарифмической гибели
В эту фазу клетки бактерий отмирают с постоянной скоростью. Ее длительность составляет около 5-и часов.
Фаза уменьшения скорости отмирания
В эту фазу оставшиеся живыми клетки бактерий переходят в состояние покоя.

Рис. 18. На рисунке отображена кривая роста бактериальной популяции.

Рис. 19. На фото колонии синегнойной палочки сине-зеленого цвета, колонии микрококков желтого цвета, колонии Bacterium prodigiosum кроваво-красного цвета и колонии Bacteroides niger черного цвета.

Рис. 20. На фото колонии бактерий. Каждая колония — потомство одной-единственной клетки. В колонии число клеток исчисляется миллионами. вырастает колония за 1 — 3 суток.
Деление магниточувствительных бактерий
В 1970-х годах были открыты бактерии, обитающие в морях, которые обладали чувством магнетизма. Магнетизм позволяет этим удивительным существам ориентироваться по линиям магнитного поля Земли и находить серу, кислород и другие, так необходимые ей вещества. Их «компас» представлен магнитосомами, которые состоят из магнита. При делении магниточувствительные бактерии делят свой компас. При этом перетяжки при делении становится явно недостаточно, поэтому бактериальная клетка сгибается и делает резкий перелом.

Рис. 21. На фото момент деления магниточувствительной бактерии.
Рост бактерий
Вначале деления бактериальной клетки две молекулы ДНК расходятся в разные концы клетки. Далее клетка делится на две равноценные части, которые отделяются друг от друга и увеличиваются до исходного размера. Скорость деления многих бактерий составляет в среднем 20 — 30 минут. В течение только одних суток одна клетка образует 72 поколения потомства.
Масса клеток в процессе роста и развития быстро поглощает питательные вещества из окружающей среды. Этому способствуют благоприятные факторы внешней среды — температурный режим, достаточное количество питательных веществ, необходимая pH среды. Для клеток аэробов необходим кислород. Для анаэробов он представляет опасность. Однако безграничное размножение бактерий в природе не происходит. Солнечный свет, сухой воздух, недостаток пищи, высокая температура окружающей среды и другие факторы губительно действуют на бактериальную клетку.

Рис. 22. На фото момент деления клетки.
Факторы роста
Для роста бактерий необходимы определенные вещества (факторы роста), часть из которых синтезируется самой клеткой, часть поступает из окружающей среды. Потребность в факторах роста у всех бактерий разная.
Потребность в факторах роста является постоянным признаком, что позволяет использовать его для идентификации бактерий, подготовке питательных сред и использовать в биотехнологии.
Факторы роста бактерий (бактериальные витамины) — химические элементы, большинством из которых являются водорастворимые витамины группы В. В эту группу входят так же гемин, холин, пуриновые и пиримидиновые основания и другие аминокислоты. При отсутствии факторов роста наступает бактериостаз.
Бактерии используют факторы роста в минимальных количествах и в неизменном виде. Ряд химических веществ этой группы входят в состав клеточных ферментов.

Рис. 23. На фото момент деления палочковидной бактерии.
Важнейшие бактериальные факторы роста
- Витамин В1 (тиамин) . Принимает участие в углеводном обмене.
- Витамин В2» (рибофлавин) . Принимает участие в окислительно-восстановительных реакциях.
- Пантотеновая кислота является составной частью кофермента А.
- Витамин В6 (пиридоксин) . Принимает участие в обмене аминокислот.
- Витамины В12 (кобаламины — вещества, содержащие кобальт). Принимают активное участие в синтезе нуклеотидов.
- Фолиевая кислота . Некоторые ее производные входят в состав ферментов, катализирующих процессы синтеза пуриновых и пиримидиновых оснований, а также некоторых аминокислот.
- Биотин . Участвует в азотистом обмене, а также катализирует синтез ненасыщенных жирных кислот.
- Витамин РР (никотиновая кислота). Участвует в окислительно-восстановительных реакциях, образовании ферментов и обмене липидов и углеводов.
- Витамин Н (парааминобензойная кислота). Является фактором роста многих бактерий, в том числе населяющих кишечник человека. Из парааминобензойной кислоты синтезируется фолиевая кислота.
- Гемин . Является составной частью некоторых ферментов, которые принимают участие в реакциях окислениях.
- Холин . Принимает участие в реакциях синтеза липидов клеточной стенки. Является поставщиком метильной группы при синтезе аминокислот.
- Пуриновые и пиримидиновые основания (аденин, гуанин, ксантин, гипоксантин, цитозин, тимин и урацил). Вещества необходимы главным образом в качестве компонентов нуклеиновых кислот.
- Аминокислоты . Эти вещества являются составляющими белков клетки.
Потребность в факторах роста некоторых бактерий
Ауксотрофы для обеспечения жизнедеятельности нуждаются в поступлении химических веществ из вне. Например, клостридии не способны синтезировать лецитин и тирозин. Стафилококки нуждаются в поступлении лецитина и аргинина. Стрептококки нуждаются в поступлении жирных кислот — компонентов фосфолипидов. Коринебактерии и шигеллы нуждаются в поступлении никотиновой кислоты. Золотистые стафилококки, пневмококки и бруцеллы нуждаются в поступлении витамина В1. Стрептококки и бациллы столбняка — в пантотеновой кислоте.
Прототрофы самостоятельно синтезируют необходимые вещества.

Рис. 24. Разные условия окружающей среды по-разному влияют на рост колоний бактерий. Слева — стабильный рост в виде медленно расширяющегося круга. Справа — быстрый рост в виде «побегов».
Изучение потребности бактерий в факторах роста позволяет ученым получать большую микробную массу, так необходимую при изготовлении антимикробных препаратов, сывороток и вакцин.
Подробно о бактерияx читай в статьях:
Размножение бактерий является механизмом повышения числа микробной популяции. Деление бактерий — основной способ размножения. После деления бактерии должны достигнуть размеров взрослых особей. Рост бактерий происходит путем быстрого поглощения питательных веществ их окружающей среды. Для роста необходимы определенные вещества (факторы роста), часть из которых синтезирует сама бактериальная клетка, часть поступает из окружающей среды.
Изучая рост и размножение бактерий, ученые постоянно открывают полезные свойства микроорганизмов, использование которых в повседневной жизни и на производстве ограничивается только их свойствами.
К царству Бактерии относятся собственно бактерии и цианобактерии.
Бактерии — это мельчайшие одноклеточные прокариотические (безъядерные) организмы.
Размеры бактерий: обычно от 0,1 до 15 мкм, но иногда достигают 30-100 мкм.
Численность видов бактерий: около 3 млрд.
Морфологические типы бактерий (в зависимости от формы тела): кокки (сферические), бациллы (прямые палочковидные), спириллы (спиралевидные), вибрионы (в виде запятой), спирохеты (извитые), колониальные формы (диплококки, стрептококки, стафилококки) и др.
Подвижность: некоторые бактерии подвижны благодаря наличию жгутиков.
В обычном состоянии бактерии неустойчивы при высушивании и воздействии прямых солнечных лучей, повышении температуры до 65-80 °С, погибают от воздействия спирта и других дезинфицирующих веществ.
Строение бактерий
Бактериальная клетка не имеет оформленного ядра, покрыта оболочкой , состоящей из плазматической мембраны, клеточной стенки и (у многих видов бактерий) внешней слизистой капсулы.
Плазматическая мембрана полупроницаема и обеспечивает избирательное поступление веществ в клетку и выделение в окружающую среду продуктов обмена веществ. Она образует складчатые впячивания внутрь цитоплазмы (мезосомы ). На мембранах мезосом находятся различные окислительно-восстановительные ферменты , участвующие в дыхании, и (у фотосинтезирующих бактерий) пигменты , участвующие в фотосинтезе. Т.е. мезосомы выполняют функции митохондрий (синтезируют АТФ), хлоропластов (осуществляют фотосинтез), комплекса Гольджи и эндоплазматической сети (накапливают и преобразуют органические вещества и осуществляют их транспорт внутри клетки и выведение за ее пределы).
Клеточная стенка — тонкая, прочная и эластичная, придает бактериальной клетке определенную форму, защищает ее содержимое от воздействия неблагоприятных факторов внешней среды и выполняет ряд других функций. Опорным каркасом клеточной стенки служит сетка из одного или несколько слоев муреина. В состав клеточной стенки бактерий не входят хитин и целлюлоза, характерные для клеток грибов и растений.
Слизистая капсула предохраняет клетку от высыхания и является ее защитным покровом, а также служит для образования колоний из отдельных клеток.
Генетический материал бактерий представлен нуклеоидом , не ограниченным мембранами и находящимся в центре клетки.
Нуклеоид (или бактериальная хромосома ) — это зона, обычно находящаяся в центре бактериальной клетки, содержащая кольцевую молекулу ДНК и не ограниченная мембранами. Молекула ДНК в нуклеоиде не связана с гистоновыми белками и прикрепляется к выросту цитоплазматической мембраны в одной точке. Нуклеоид является носителем генетической информации и контролирует нормальный ход всех внутриклеточных процессов.
Молекула ДНК у бактерий имеет до 5 000 000 пар нуклеотидов ; но суммарное содержание ДНК в одной бактериальной клетке значительно меньше, чем в ядерной (эукариотической).
Цитоплазма бактериальной клетки представляет собой смесь белков, жиров, углеводов, других органических соединений, минеральных веществ и воды и имеет зернистый вид. В ней содержится до 20 тысяч рибосом класса 70S (медленно осаждаемых), на которых синтезируются белки. В цитоплазме бактерий также содержатся многочисленные включения — гранулы запасаемых веществ. У некоторых бактерий в цитоплазме имеются плазмиды — небольшие кольцевые молекулы ДНК, участвующие в обмене генетической информацией между различными бактериальными клетками.
В клетках бактерий отсутствуют митохондрии, лизосомы, комплекс Гольджи и другие органеллы, однако в них хорошо развиты мембранные структуры в виде канальцев, пузырьков и тила-коидов, часто содержащих ферменты и пигменты и являющихся аналогами многих органелл эукариотической клетки.
Жгутики — это органоиды движения бактерий, состоящие из собранных в спираль глобул особого белка — флагеллина . Они берут свое начало под цитоплазматической мембраной, закрепляясь там с помощью пары дисков. Количество жгутиков у бактерии — от I до 50. У одних бактерий жгутики расположены только на одном конце клетки, у других — на двух или по всей поверхности. Способ расположения жгутиков является характерным признаком при классификации подвижных бактерий.
У некоторых безжгутиковых водных и почвенных бактерий в цитоплазме имеются газовые вакуоли , позволяющих погружаться в толщу воды, подниматься на ее поверхность или передвигаться в капиллярах почвы.
Классификация бактерий
❖ Классификация бактерий по типу питания (ассимиляции):
■ автотрофные,
■ гетеротрофные.
♦Автотрофные бактерии сами синтезируют нужные им органические вещества из неорганических.
■ В зависимости от способа получения энергии, необходимой для этого синтеза, автотрофные бактерии подразделяются на фотосинтезирующие и хемосинтезирующие . Фотосинтезирующие бактерии (например, зеленые и пурпурные) осуществляют фотосинтез органических веществ, используя световую (солнечную) энергию.
В клетках фотосинтезирующих бактерий (в отличие от клеток растений) нет пластид, а фотосинтезирующие пигменты (бактерио-хлорофиллы ) находятся в тилакоидах, образующихся в результате выпячивания цитоплазматической мембраны. По своей структуре бактериохлорофиллы подобны хлорофиллам растений и отличаются от них природой белковых цепей.
Хемосинтезирующие бактерии получают нужную для синтеза энергию от экзотермических реакций окисления неорганических веществ (молекулярного водорода, сероводорода, аммиака, закиси железа и др.). ‘
❖ Гетеротрофные бактерии (их большинство) используют в пищу готовые органические вещества, которые служат этим бактериям источником энергии и атомов углерода.
■ В зависимости от источника пищи гетеротрофные бактерии подразделяются на сапротрофы и симбионты .
Сапротрофы извлекают органические вещества из разлагающихся мертвых остатков организмов (бактерии гниения , получающие энергию от расщепления азотсодержащих соединений), выделений живых организмов (бактерии брожения , получающие энергию от расщепления углеродсодержащих соединений).
Симбионты поглощают органические вещества тела хозяина (растения, животного или человека), в котором они живут. При этом симбионты или:
■ продуцируют вещества, необходимые организму хозяина (пример: клубеньковые азотфиксирующие бактерии, поселяющиеся на корнях бобовых растений и находящиеся с ними во взаимовыгодном сосуществовании), или
❖ Классификация бактерий по типу диссимиляции
(потребности в кислороде для высвобождения энергии, запасенной в молекулярных связях):
■ аэробные,
■ анаэробные,
■ факультативные.
Аэробные бактерии (туберкулезная палочка, гнилостные бактерии) живут только в кислородной среде (в верхних слоях почвы, в воздухе) и получают энергию путем окисления органических соединений до воды и диоксида углерода.
Анаэробные бактерии (бактерии желудочно-кишечного тракта, столбнячная палочка, возбудители гангрены, палочка ботулизма и др.) обитают в бескислородных средах и получают энергию в процессе реакций гликолиза и брожения.
Факультативные бактерии могут обитать как в кислородных, так и в бескислородных средах (пример: молочнокислая бактерия).
Размножение бактерий
❖ Тип размножения бактерий — бесполый . Бактериальная клетка начинает размножаться, попав в благоприятные условия и достигнув определенного размера.
❖ Формы (способы) размножения бактерий:
■ делением клетки надвое,
■ почкованием (встречается как исключение),
■ спорообразованием.
Размножение делением клетки надвое: сначала путем репликации ДНК удваивается генетический материал клетки. После этого белки, прикрепляющие молекулы ДНК к выростам цитоплазматической мембраны, разделяют (растаскивают) дочерние молекулы ДНК и происходит оформление обособленных бактериальных хромосом (нуклеоидов ). Затем клетка удлиняется, и в ней постепенно образуется поперечная перегородка. Наконец, две дочерние клетки расходятся. Деления клеток происходят примерно через каждые 15-20 минут.
Спорообразование свойственно некоторым бактериям при наступлении неблагоприятных условий. При этом в бактериальной клетке значительно уменьшается количество свободной воды, снижается ферментативная активность, цитоплазма сжимается, а клетка покрывается очень плотной оболочкой. Споры бактерий устойчивы к различным воздействиям (выдерживают длительное высыхание, нагревание свыше 100 °С и охлаждение примерно до -200 °С) и сохраняют жизнеспособность в течение длительного времени. При попадании в благоприятные условия споры набухают и прорастают, образуя новую вегетативную клетку бактерий.
♦ Виды спор бактерий:
■ микроцисты
(образуются из целой клетки),
■ эндогенные
(образуются внутри клетки).
Циста — временная форма существования многих одноклеточных и ряда простейших многоклеточных организмов, характеризующаяся наличием защитной оболочки. Позволяет перенести неблагоприятные условия или предохраняет клетку в период ее деления.
❖ Формы полового процесса у бактерий:
■ трансформация,
■ конъюгация,
■ трансдукция.
Трансформация осуществляется при попадании фрагментов ДНК разрушенных клеток одной культуры бактерий в живую культуру другой бактерии. Эти фрагменты ДНК могут поглощаться клеткой-реципиентом и встраиваться в ее нуклеоид.
При конъюгации перенос участка ДНК от донора (выполняющего мужские функции) к клетке-реципиенту осуществляется при непосредственном контакте через половую фимбрию (тонкую белковую трубочку), которая формируется у клетки-донора. После этого клетки разъединяются. При конъюгации очень часто наблюдается передача не всей молекулы ДНК, а только ее фрагментов.
При трансдукции небольшой фрагмент ДНК переносится от одной клетки к другой бактериофагами .
Значение бактерий
❖ Положительное значение:
■ они участвуют в круговороте веществ и являются конечным звеном всех цепей питания;
■ являются редуцентами в биогеоценозе (разлагают и минерализуют экскременты и органические остатки);
■ участвуют в процессе почвообразования;
■ служат источником азота для бобовых растений;
■ принимают участие в образовании торфа, каменного угля, железной руды, других полезных ископаемых;
■ участвуют в биохимических процессах пищеварения животных и человека;
■ применяются в пищевой промышленности (для консервирования, получения молочнокислых продуктов и т.д.);
■ используются в микробиологической и химической промышленности (для получения спиртов, ацетона, сахаров, органических кислот и других химических соединений),
■ используются в фармацевтической промышленности для получения антибиотиков, вакцин, витаминов, аминокислот, ферментов и других биологически активных веществ;
■ применяются в процессах обработки льна, дубления кож и т.д.;
■ являются удобным объектом для генной инженерии;
■ применяются для борьбы с вредителями сельского хозяйства.
Дифтерия вызывается дифтерийной палочкой , поражающей верхние дыхательные пути. Токсин, выделяемый этими бактериями, разносится кровью и воздействует на сердце. Способ борьбы — прививка неактивным токсином.
Тиф: возбудитель — бактерии риккетсии , их переносчик -вши. При заболевании поражаются стенки кровеносных сосудов и образуются тромбы. Возможна прививка с помощью убитых бактерий, а также лечение антибиотиками тетрациклинового ряда.
Туберкулез: возбудитель — туберкулезная палочка , поражающая легкие и кости. Заражение происходит воздушно-капельным путем, а также через молоко больных животных. Профилактика -вакцинацией; лечение производится специальными препаратами.
Сифилис: возбудитель — спирохета рода трепонема . Сначала поражаются половые органы, затем глаза, кости, суставы, кожа, центральная нервная система. Передается при половом контакте. Лечение — антибиотиками и специальными препаратами.
Холера вызывается холерным вибрионом , в результате жизнедеятельности которого выделяется токсин, поражающий слизистую кишечника. Заражение происходит при употреблении в пищу грязных продуктов питания и воды. Для лечения применяются антибиотики тетрациклинового ряда.
■ Токсины — ядовитые продукты жизнедеятельности бактерий, которые, как правило, или сами являются поражающими факторами, или угнетают защитные силы организма, усиливая патогенное действие возбудителей болезни.
Методы борьбы с бактериями
❖ Методы борьбы с гнилостными бактериями:
■ высушивание плодов, грибов, мяса, рыбы, зерна;
■ охлаждение и замораживание продуктов;
■ маринование продуктов в уксусной кислоте;
■ создание высокой концентрации сахара (например, при изготовлении варенья), что вызывает плазмолиз в клетках бактерий и нарушает их жизнедеятельность;
■ консервирование (засолка).
❖ Другие методы борьбы с бактериями, в том числе болезнетворными:
■ дезинфекция (обеззараживание) — уничтожение болезнетворных микроорганизмов специальными химическими веществами (хлорной известью, хлорамином, раствором йода, этиловым спиртом и др.);
■ пастеризация — уничтожение бактерий в пищевых продуктах нагреванием до температуры 65-70 °С в течение 15-30 мин;
■ стерилизация — уничтожение бактерий с помощью ультрафиолетового излучения, химикатов или кипячения в автоклавах при температуре 120-130 °С и повышенном давлении;
■ соблюдение гигиены;
■ профилактические прививки.
Цианобактерии
Цианобактерии (или сине-зеленые водоросли ) — группа микроскопических фототрофных одноклеточных, колониальных и многоклеточных (нитчатых) прокариотических организмов.
■ Цианобактерии осуществляют обычный двухфазный (со световой и темновой фазами) кислородный фотосинтез.
❖ Распространение: в пресных и соленых водоемах (входят в состав планктона и бентоса ), на поверхности почвы, на скалах; могут вступать в симбиоз с грибами (образуя лишайники), протистами, водорослями, мхами.
Планктон — совокупность организмов (бактерий, микроскопических водорослей, животных и их личинок), населяющих толщу воды и пассивно переносимых течением.
Бентос — совокупность организмов, обитающих в грунте и на поверхности дна водоема.
❖ Строение — сходное с бактериями: клетки безъядерные , имеют толстые многослойные стенки , состоящие из полисахаридов, пектиновых веществ и целлюлозы; часто покрыты слизистым чехлом. В цитоплазме расположены мембранные фотосинтезирующие структуры и пигменты , хлорофиллы, каротиноиды, фикоэритрин и др. (благодаря их разнообразию цианобактерии могут поглощать свет различных длин волн), а также нуклеоид, рибосомы, включения запасного вещества —гликоген а, а также (у некоторых видов) газовые вакуоли , наполненные азотом и регулирующие плавучесть клетки. У ряда нитчатых форм цианобактерий имеются специализированные клетки с сильно утолщенными бесцветными оболочками — гетероцисты, участвующие в фиксации азота и размножении.
❖ Размножение: бесполое, делением клетки надвое; колониальные и нитчатые цианобактерии — распадом колоний или нитей.
♦ Значение бактерий:
■ обогащают воду кислородом, а почву — органикой и азотом;
■ очищают воду, минерализуя продукты гниения;
■ являются кормом для зоопланктона и рыб;
■ используются для получения ряда ценных веществ (аминокислот, пигментов, витамина В 12 и др.), вырабатываемых ими в процессе жизнедеятельности;
■ отдельные виды (спирулина, носток) используются в пищу;
■ (отрицательное) вызывают «цветение» воды в период массового размножения, обычно сопровождающегося гибелью (из-за недостатка пищи) и гниением большинства дочерних особей, что делает воду непригодной для питья и вызывает гибель рыбы.
22. Бактериальное ядро. Виды деления бактериальной клетки. Процесс деления.
Виды деления:
1. Равновеликое бинарное поперечное деление , приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. При равновеликом бинарном делении материнская клетка, делясь, дает начало двум дочерним клеткам и сама, таким образом, исчезает.
2. Неравновеликое бинарное деление, или почкование . При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново. В процессе почкования симметрия наблюдается в отношении только продольной оси. При почковании материнская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя.
3. Размножение путем множественного деления , характерное для одной группы одноклеточных цианобактерий, в результате образуются мелкие клетки, получивших название баеоцитов (греч. bae – маленькая, cyto – клетка), число которых у разных видов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки. В основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
23. Бактериальное ядро. Формы обмена генетической информацией у бактерий. Изменчивость бактерий.
Формы обмена генетическим материалом у бактерий:
1. по горизонтали
* трансформация – перенос генетического материала, заключающийся в том, что бактерия-реципиент захватывает (поглощает) из внешней среды фрагменты чужеродной ДНК.
А) Индуцированная (искусственно получаемая) трансформация происходит при добавлении к культуре бактерий очищенной ДНК, полученной из культур тех бактерий, генетические признаки которых стремятся передать исследуемой культуре.
Б) Спонтанная трансформация происходит в естественных условиях и проявляется в возникновении рекомбинантов при смешивании генетически различающихся клеток. Она протекает за счет ДНК, выделяющейся клетками в окружающую среду вследствие их лизиса или в результате активного выделения ДНК жизнеспособными клетками-донорами.
* сексдукция
* трансфекция – вариант трансформации бактериальных клеток, лишенных клеточной стенки, осуществляемый вирусной (фаговой) нуклеиновой кислотой. С помощью трансфекции удается вызвать у таких бактерий (без клеточной стенки) вирусную инфекцию. Трансфекцию можно осуществить и с другими (не бактериальными) клетками, если ввести в них чужеродную ДНК, способную рекомбинировать с ДНК этих клеток, или воспроизводить вирионы, или самостоятельно реплицироваться.
* конъюгация – процесс обмена генетическим материалом (хромосомным и плазмидным), осуществляемый при непосредственном контакте клеток донора и реципиента. Этот процесс контролируется только конъюгативными плазмидами, имеющими совокупность генов, называемую tra-опероном (tra – от англ., transfer – перенос).
Этот оперон контролирует синтез аппарата переноса, конъюгативную репликацию и явление поверхностного исключения. Аппаратом переноса являются специальные донорные ворсинки, с помощью которых устанавливается контакт между конъюгирующими клетками. Донорные ворсинки представляют собой длинные (1-20 мкм) тонкие трубчатые структуры белковой природы с внутренним диаметром около 3 нм.
установление контакта между донором и реципиентом
протаскивание нити ДНК от донора к реципиенту
достройка перенесенной нити ДНК комплементарной ей нитью в реципиентной клетке
рекомбинация между переданной хромосомой (ее фрагментами) и хромосомой клетки-реципиента
размножение мерозиготы
образование клеток, несущих признаки донора и реципиента
Конъюгативная репликация переносимой нити хромосомной или плазмидной ДНК осуществляется также под контролем плазмидных генов. Классическим примером конъюгативной плазмиды является половой фактор, или F-плазмида (от англ. fertility – плодовитость). F-плазмида может находиться как в автономном состоянии, так и интегрироваться в хромосому клетки. Находясь в автономном состоянии, она контролирует только собственный перенос, при котором Р~-клетка (клетка, лишенная F-плазмиды) превращается в Р+-клетку (клетку, содержащую F-плазмиду). F-плазмида может интегрироваться в определенные участки бактериальной хромосомы, в этом случае она станет контролировать конъюгативный перенос хромосомы клетки.
Таким образом, конъюгация начинается с установления контакта между донором и реципиентом с помощью донорной ворсинки. Последняя смыкается с рецептором клеточной мембраны клетки-реципиента. Нередко такой контакт устанавливается не только между двумя клетками, а между многими клетками, образуя агрегаты спаривания. Предполагают, что нить ДНК в процессе конъюгации протаскивается через канал донорной ворсинки. Поскольку донорный мостик является непрочным, процесс конъюгации может в любой момент прерваться. Поэтому при конъюгации может переноситься или часть хромосомы, или, реже, – полная хромосома. С помощью F-плазмид частота переноса генов между бактериями существенно возрастает.
* трансдукция - перенос генетического материала от клетки-донора клетке-реципиенту с помощью бактериофагов. Различают трансдукцию неспецифическую и специфическую.
А) Неспецифическая трансдукция - случайный перенос фрагментов ДНК от одной бактериальной клетки к другой.
Б) Специфическая трансдукция осуществляется только умеренными фагами, способными включаться в строго определенные участки хромосомы бактериальной клетки и переносить определенные гены.
Молекулярные механизмы изменчивости бактерий
Бактерии в силу относительной простоты их организации и короткого срока жизни подвергаются изменчивости быстрее, чем многие другие организмы. В основе их изменчивости лежат мутации и генетические рекомбинации, особенно протекающие с участием транспонируемых элементов.
*Мутации – изменения в генотипе, которые стабильно наследуются. Мутации могут быть спонтанными или индуцированными.
а) Спонтанные мутации возникают без каких-либо специальных воздействий, они происходят в результате ошибок при репликации и репарации. Средняя частота спонтанных мутаций составляет около 1 106 (один мутант на 1 млн. клеток).
б) Индуцированные мутации происходят с гораздо большей частотой, они возникают в результате воздействия различных мутагенов – физических и химических факторов, повреждающих ДНК: ионизирующая радиация, УФ облучение, различные аналоги оснований ДНК, алкилирующие соединения, акридины, антибиотики
в) Точечные мутации могут быть обусловлены: заменой оснований, выпадением (делецией) основания, появлением дополнительного основания (вставки). Точечные мутации могут иметь три последствия:
1) замена одного кодона на другой, а стало быть, одной аминокислоты на другую;
2) сдвиг рамки считывания, что приведет к изменению целой серии последовательностей аминокислотных остатков;
3)возникновение «бессмысленного» кодона, что приведет к прекращению трансляции в данной точке
синтез белка может быть полностью заблокирован. Будет синтезироваться измененный белок
Все это приведет либо к утрате какого-то фенотипического признака у мутанта, либо, реже, к появлению у него нового признака.
Нарушение генома может быть следствием:
*протяженных делеций
*инверсии (поворот сегмента хромосомы на 180°)
*транслокации (перемещение участка хромосомы из одной позиции в другую)
Все это также будет приводить к изменению и нарушению различных функций клетки (организма).
Большая роль в изменчивости бактерий и других организмов принадлежит так называемым транспонируемым генетическим элементам, то есть генетическим структурам, способным в интактной форме перемещаться внутри данного генома или переходить от одного генома к другому, например от плазмидного генома к бактериальному и наоборот. Различают три класса транспонируемых элементов: IS-элементы, транспозоны и эписомы.
#Вставочные последовательности (от англ, insertion sequence), имеют обычно размеры, не превышающие 2 тыс. пар оснований, или 2 к.б. (килобаза – тысяча пар оснований). IS-элементы несут только один ген, кодирующий белок транспозазу, с помощью которой IS-элементы встраиваются в различные участки хромосомы. Их обозначают цифрами: IS1, IS2, IS3 и т. д.
#Транспозоны представляют собой более крупные сегменты ДНК, фланкированные инвертированными IS-элементами. Способны встраиваться в различные участки хромосомы или переходить из одного генома в другой, т. е. ведут себя как IS-элементы. Помимо генов, обеспечивающих их перемещение, они содержат и другие гены, например гены лекарственной устойчивости. Транспозоны обнаружены в геномах плазмид, вирусов, прокариот и эукариот и их, как и IS-элементы, обозначают порядковым номером: Tп1, Тп2, ТпЗ и т. д.
# К эписомам относятся еще более крупные и сложные саморегулирующиеся системы, содержащие IS-элементы и транспозоны и способные реплицироваться в любом из двух своих альтернативных состояний – автономном или интегрированном – в хромосому клетки-хозяина. К эписомам относят различные умеренные лизогенные фаги; они отличаются от всех других транспонируемых элементов наличием собственной белковой оболочки и более сложным циклом репродукции. Собственно эписомы – это вирусы, обладающие, подобно другим транспонируемым элементам, способностью в интактной форме переходить из одного генома в другой.
Обычно деление бактериальных клеток описывается как "бинарное": после удвоения нуклеоиды, связанные с плазматической мембраной, расходятся за счет растяжения мембраны между нуклеоидами, а затем образуется перетяжка или септа, делящая клетку надвое. Этот тип деления приводит к очень точному распределению генетического материала, практически без ошибок (менее 0,03 % дефектных клеток). Напомним, что ядерный аппарат бактерий, нуклеоид, представляет собой циклическую гигантскую (1,6 мм) молекулу ДНК, образующую многочисленные петлевые домены в состоянии сверхспирализации, порядок укладки петлевых доменов не известен.
Среднее время между делениями бактериальных клеток составляет 20-30 мин. А это период должен произойти целый ряд событий: репликация ДНК нуклеоида, сегрегация, отделение сестринских нуклеоидов, их дальнейшее расхождение, цитотомия за счет образования септы, делящей исходную клетку ровно пополам.
Весь ряд этих процессов находится под интенсивным вниманием исследователей последних лет, в результате были получены важные и неожиданные наблюдения. Так оказалось, что в начале синтеза ДНК, который начинается с точки репликации (origin), обе растущие молекулы ДНК изначально остаются связанными с плазматической мембраной. Одновременно с синтезом ДНК происходит процесс снятия сверхспирализации как старых, так и реплицирующихся петлевых доменов за счет целого ряда ферментов (топоизомеразы, гиразы, лигазы и др), что приводит к физическому обособлению двух дочерних (или сестринских) хромосом-нуклеоидов, которые еще находятся в тесном контакте друг с другом. После такой сегрегации нуклеоидов происходит их расхождение от центра клетки, от места их бывшего расположения. Причем это расхождение очень точное: на четверть длины клетки в двух противоположных направлениях. В результате этого в клетке располагаются два новых нуклеоида. Каков механизм этого расхождения? Делались предположения (Деламатер, 1953), что деление бактериальных клеток аналогично митозу эукариот, однако данных в пользу этого предположения долгое время не появлялось.
Новые сведения о механизмах деления бактериальных клеток были получены при изучении мутантов, в которых происходили нарушения клеточного деления.
Было обнаружено, что в процессе расхождения нуклеоидов принимают участие несколько групп специальных белков. Один из них, белок Muk В, представляет собой гигантский гомодимер (мол.масса около 180 кДа, длина 60 нм), состоящий из центрального спирального участка, и концевых глобулярных участков, напоминающий по структуре нитевидные белки эукариот (цепь миозина II, кинезина). На N-конце Muk В связывается с ГТФ и АТФ, а на С-конце — с молекулой ДНК. Эти свойства Muk В дают основания считать его моторным белком, участвующим в расхождении нуклеоидов. Мутации этого белка приводят к нарушениям расхождения нуклеоидов: в мутантной популяции появляется большое количество безъядерных клеток.
Кроме белка Muk В в расхождении нуклеоидов, по-видимому, участвуют пучки фибрилл, содержащих белок Caf A, который может связываться с тяжелыми цепями миозина, подобно актину.
Образование перетяжки, или септы также в общих чертах напоминает цитотомию животных клеток. В данном случае в образовании септ принимают участие белки семейства Fts (фибриллярные термочувствительные). Это группа из нескольких белков, среди которых наиболее изучен белок FtsZ. Этот белок сходен у большинства бактерий, архибактерий, обнаружен в микоплазмах и хлоропластах. Это глобулярный белок, сходный по своей аминокислотной последовательности с тубулином. При взаимодействии с ГТФ in vitro он способен образовывать длинные нитчатые протофиламенты. В интерфазе FtsZ диффузно локализуется в цитоплазме, его количество очень велико (5-20 тыс. мономеров на клетку). Во время деления клетки весь этот белок локализуется в зоне септы, образуя сократимое кольцо, очень напоминающее акто-миозиновое кольцо при делении клеток животного происхождения.
22. Бактериальное ядро. Виды деления бактериальной клетки. Процесс деления.
Мутации по этому белку приводят к прекращению деления клеток: возникают длинные клетки, содержащие множество нуклеоидов. Эти наблюдения показывают прямую зависимость деления бактериальных клеток от наличия Fts-белков.
Относительно механизма образования септ существует несколько гипотез, постулирующих сокращение кольца в зоне септы, приводящее к разделению исходной клетки надвое. По одной из них протофиламенты должны скользить один относительно другого с помощью неизвестных еще моторных белков, по другой — сокращение диаметра септы может происходить за счет деполимеризации заякоренных на плазматической мембране FtsZ.
Фазы размножения культуры бактерий в стационарных условиях
Последняя фаза роста - стационарная фаза, которая вызвана истощением питательных веществ. Клетки сокращают свою метаболическую деятельность и потребляют несущественные клеточные белки. Стационарная фаза - это переход от быстрого роста к стрессовому состоянию, которое характеризуется увеличением экспрессии генов, которые принимают участие в ремонте ДНК и антиоксидантном метаболизме.
При размножении бактерий не в проточных, а в стационарных условиях происходит изменение питательной среды и накопление в ней продуктов жизнедеятельности бактерий, вследствие чего меняются и их физиологические особенности. Так, молодые клетки Clostridium acetobutylicum не способны образовывать ацетон; это свойство они приобретают в более старой культуре. Если спороносных бактерий выращивать в условиях проточной культуры, они будут делиться, но не будут давать спор. При выращивании бактерий на плотных питательных средах они образуют скопления клеток разных размеров, формы, цвета, называемые колониями.
Штамм бактерий, чистая культура
Особенности полового размножения
Типы бесполого размножения, термины
Глоксиния, виды, заболевания
Влияние гормонов на организм
Этапы гликолиза
Пентозомонофосфатный путь окисления глюкозы
Цикл Кребса
Деление бактерий
Деление бактерий наступает в результате формирования межклеточной перегородки, которое происходит следующим образом. В том участке ЦМ, с которым связана с помощью особого рецептора молекула ДНК (хромосома, плазмида), происходят события, инициирующие процесс репликации, в результате которого вновь образующаяся дочерняя молекула ДНК прикрепляется также к рецептору на ЦМ.
Область последней между двумя рецепторами, к одному из которых прикреплена родительская, а к другому - дочерняя ДНК, начинает удлиняться, в результате этого расстояние между ними все время увеличивается в течение времени. По завершении процесса репликации строго по экватору между отделившимися друг от друга хромосомами начинает формироваться межклеточная перегородка путем встречной инвагинации (врастания навстречу друг к другу) ЦМ и связанной с ней области клеточной стенки.
В результате слияния инвагинирующих участков ЦМ и КС образуется межклеточная перегородка, и родительская клетка разделяется на две дочерние клетки равной длины, функцию аппарата митоза у бактерий выполняет ЦМ путем своего удлинения, которое раздвигает хромосомы (и плазмиды) таким образом, что они оказываются по ту и другую стороны формирующейся межклеточной перегородки в равных соотношениях.
Результатов нарушения генетического контроля клеточного деления может быть по крайней мере два. Если формирования межклеточной перегородки не происходит, возникают длинные нитевидные формы. Однако при восстановлении нарушенного механизма такого контроля нити делятся на фрагменты, равные по длине нормальным клеткам. В некоторых случаях нарушение контрольных механизмов приводит к тому, что вместо одной межклеточной перегородки, формирующейся по экватору, происходит образование одной или двух перегородок, каждая из которых локализована ближе к своему полюсу.
Поскольку в этом случае формирование перегородки не связано с сегрегацией хромосом, образуются так называемые мини-клетки, лишенные хромосом, которые остаются в родительской клетке. Мини-клетки могут осуществлять различные биохимические процессы, поскольку они содержат ферменты, но они не способны к размножению, так как лишены хромосом.
Помимо мини-клеток вследствие различных неблагоприятных воздействий из бактерий могут образовываться так называемые нанно-клетки, т. е. мельчайшие клетки размером 0,2-0,3 мкм. Их описывали под различными названиями: фильтрующиеся формы бактерий, элементарные тельца, ультрамикробактерии.
Деление бактериальной клетки
Чаще всего они образуются при L-трансформации бактерий.
Поскольку размеры таких клеток удобнее выражать в нанометрах, а не в долях микрометра, их стали называть наноклетками. Образование наноклеток - универсальная ответная реакция бактерий на неблагоприятные условия существования.
Не нашли подходящую информацию? Не беда! Воспользуйтесь поиском на сайте в верхнем правом углу.
1. Какие способы деления характерны для клеток эукариот? Для прокариотических клеток?
Митоз, амитоз, простое бинарное деление, мейоз.
Для клеток эукариот характерны следующие способы деления: митоз, амитоз, мейоз.
Для прокариотических клеток характерно простое бинарное деление.
2. Что представляет собой простое бинарное деление?
Простое бинарное деление характерно только для клеток прокариот. Бактериальные клетки содержат одну хромосому – кольцевую молекулу ДНК. Перед делением клетки происходит репликация и образуются две одинаковые молекулы ДНК, каждая из них прикреплена к цитоплазматической мембране. Во время деления плазмалемма врастает между двумя молекулами ДНК таким образом, что в итоге разделяет клетку надвое. В каждой образовавшейся клетке оказывается по одной идентичной молекуле ДНК.
3. Что такое митоз? Охарактеризуйте фазы митоза.
Митоз – основной способ деления эукариотических клеток, в результате которого из одной материнской клетки образуются две дочерние с таким же набором хромосом. Для удобства митоз подразделяют на четыре фазы:
● Профаза. В клетке увеличивается объём ядра, начинает спирализоваться хроматин, в результате чего формируются хромосомы. Каждая хромосома состоит из двух сестринских хроматид, соединённых в области центромеры (в диплоидной клетке – набор 2n4c). Растворяются ядрышки, распадается ядерная оболочка. Хромосомы оказываются в гиалоплазме и располагаются в ней беспорядочно (хаотически). Центриоли попарно расходятся к полюсам клетки, где инициируют образование микротрубочек веретена деления. Часть нитей веретена деления идёт от полюса к полюсу, другие нити прикрепляются к центромерам хромосом и способствуют их перемещению в экваториальную плоскость клетки. В клетках большинства растений центриоли отсутствуют. В этом случае центрами образования микротрубочек веретена деления являются особые структуры, состоящие из мелких вакуолей.
● Метафаза. Завершается формирование веретена деления. Хромосомы достигают максимальной спирализации и располагаются упорядоченно в экваториальной плоскости клетки. Образуется так называемая метафазная пластинка, состоящая из двухроматидных хромосом.
● Анафаза. Нити веретена деления укорачиваются, в результате чего сестринские хроматиды каждой хромосомы отделяются друг от друга и растягиваются к противоположным полюсам клетки. С этого момента разошедшиеся хроматиды называются дочерними хромосомами. У полюсов клетки оказывается одинаковый генетический материал (у каждого полюса – 2n2c).
● Телофаза. Дочерние хромосомы деспирализуются (раскручиваются) у полюсов клетки с образованием хроматина. Вокруг ядерного материала каждого полюса формируются ядерные оболочки. В двух образовавшихся ядрах возникают ядрышки. Нити веретена деления разрушаются. На этом деление ядра заканчивается, и начинается разделение клетки надвое. У клеток животных в экваториальной плоскости возникает кольцевая перетяжка, которая углубляется до тех пор, пока не произойдёт разделение двух дочерних клеток. Клетки растений не могут делиться перетяжкой, т.к. имеют жёсткую клеточную стенку. В экваториальной плоскости растительной клетки из содержимого пузырьков комплекса Гольджи образуется так называемая срединная пластинка, которая и разделяет две дочерние клетки.
4. Благодаря чему дочерние клетки в результате митоза получают идентичную наследственную информацию? В чём заключается биологическое значение митоза?
В метафазе в экваториальной плоскости клетки находятся двухроматидные хромосомы. Молекулы ДНК в составе сестринских хроматид идентичны друг другу, т.к. образовались в результате репликации исходной материнской молекулы ДНК (это произошло в S-периоде интерфазы, предшествующей митозу).
В анафазе с помощью нитей веретена деления сестринские хроматиды каждой хромосомы отделяются друг от друга и растягиваются к противоположным полюсам клетки. Таким образом, у двух полюсов клетки оказывается одинаковый генетический материал (2n2c у каждого полюса), который по завершении митоза становится генетическим материалом двух дочерних клеток.
Биологическое значение митоза заключается в том, что он обеспечивает передачу наследственных признаков и свойств в ряду поколений клеток. Это необходимо для нормального развития многоклеточного организма. Благодаря точному и равномерному распределению хромосом при митозе все клетки организма генетически идентичны. Митоз обусловливает рост и развитие организмов, восстановление повреждённых тканей и органов (регенерацию). Митотическое деление клеток лежит в основе бесполого размножения многих организмов.
5. Количество хромосом - n, хроматид - с. Каким будет соотношение n и с для соматических клеток человека в следующих периодах интерфазы и митоза. Установите соответствие:
1) В G1-периоде каждая хромосома состоит из одной хроматиды, т.е. соматические клетки содержат набор 2n2с, что для человека составляет 46 хромосом, 46 хроматид.
2) В G2-периоде каждая хромосома состоит из двух хроматид, т.е. соматические клетки содержат набор 2n4с (46 хромосом, 92 хроматиды).
3) В профазе митоза набор хромосом и хроматид – 2n4c, (46 хромосом, 92 хроматиды).
4) В метафазе митоза набор хромосом и хроматид – 2n4c (46 хромосом, 92 хроматиды).
5) В конце анафазы митоза вследствие отделения сестринских хроматид друг от друга и их расхождения к противоположным полюсам клетки, у каждого полюса оказывается набор 2n2с (46 хромосом, 46 хроматид).
6) В конце телофазы митоза формируются две дочерние клетки, каждая содержит набор 2n2c (46 хромосом, 46 хроматид).
Ответ: 1 – В, 2 – Г, 3 – Г, 4 – Г, 5 – В, 6 – В.
6. Чем амитоз отличается от митоза?
Деление прокариотических клеток
Как вы думаете, почему амитоз называют прямым делением клетки, а митоз - непрямым?
В отличие от митоза при амитозе:
● Происходит деление ядра перетяжкой без спирализации хроматина и образования веретена деления, отсутствуют все четыре фазы, характерные для митоза.
● Наследственный материал распределяется между дочерними ядрами неравномерно, случайным образом.
● Часто наблюдается только деление ядра без дальнейшего разделения клетки на две дочерние. В этом случае возникают двуядерные и даже многоядерные клетки.
● Затрачивается меньше энергии.
Митоз называют непрямым делением, т.к. по сравнению с амитозом он представляет собой достаточно сложный и точный процесс, состоящий из четырёх фаз и требующий предварительной подготовки (репликации, удвоения центриолей, запасания энергии, синтеза специальных белков и т.д.). При прямом (т.е. простом, примитивном) делении – амитозе ядро клетки без какой-либо специальной подготовки быстро делится перетяжкой, и наследственный материал случайным образом распределяется между дочерними ядрами.
7. В ядре неделящейся клетки наследственный материал (ДНК) находится в виде аморфного рассредоточенного вещества - хроматина. Перед делением хроматин спирализуется и образует компактные структуры - хромосомы, а после деления возвращается в исходное состояние. Для чего клетки совершают такие сложные видоизменения своего наследственного материала?
ДНК в составе аморфного и рассредоточенного хроматина при делении было бы невозможно точно и равномерно распределить между дочерними клетками (именно такая картина и наблюдается при амитозе – наследственный материал распределяется неравномерно, случайным образом).
С другой стороны, если бы клеточная ДНК всегда находилась в компактизированном состоянии (т.е. в составе спирализованных хромосом), с неё было бы невозможно считывать всю необходимую информацию.
Поэтому клетка в начале деления переводит ДНК в максимально компактное состояние, а после завершения деления возвращает в исходное, удобное для считывания.
8*. Установлено, что у дневных животных максимальная митотическая активность клеток наблюдается вечером, а минимальная - днём. У животных, ведущих ночной образ жизни, клетки наиболее интенсивно делятся утром, ночью же митотическая активность ослаблена. Как вы думаете, с чем это связано?
Дневные животные активны в светлое время суток. Днём они затрачивают много энергии на передвижение и поиск пищи, при этом их клетки быстрее "изнашиваются" и чаще погибают. Вечером, когда организм переварил пищу, усвоил питательные вещества и накопил достаточное количество энергии, активизируются процессы регенерации и, прежде всего, митоз. Соответственно, у ночных животных максимальная митотическая активность клеток наблюдается утром, когда их организм отдыхает после активного ночного периода.
* Задания, отмеченные звёздочкой, предполагают выдвижение учащимися различных гипотез. Поэтому при выставлении отметки учителю следует ориентироваться не только на ответ, приведённый здесь, а принимать во внимание каждую гипотезу, оценивая биологическое мышление учащихся, логику их рассуждений, оригинальность идей и т. д. После этого целесообразно ознакомить учащихся с приведённым ответом.
Дашков М.Л.
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки | 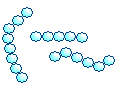 | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.